Механизм передвижеия воды из корня в надземные органы
Механизм передвижеия воды из корня в надземные органы. Верхний и нижний концевые двигатели.
Т. к. вода поглощается в осн. корневой системой, возникает необходимость ее передвижения по растению. Этот процесс составляет т. н. восходящий ток. Следует отметить, что это название отражает характер передвижения и его локализацию в растении. Он проходит по мертвым тканям стебля или черешка — сосудам у покрытосеменных и трахеидам у голосеменных. Однако эта локализация не абсолютна: вода способна перемещаться и по флоэмной системе. Передвижение по сосудам идет с большей скоростью (20 см/ч), чем по трахеидам (только 5 см/ч).
Если принять во внимание всю протяженность пути восходящего тока, то его можно будет разбить на 2 не равных по длине участка: Мертвые гистологические элементы в середине проводящего пути — сосуды или трахеиды. Протяженность значительна, но вода проходит по нему сравнительно легко, так как двигается пассивно по мертвым элементам, не испытывая с их стороны значительного сопротивления. Живые клетки корня и листа, находящиеся в начале и в конце пути передвижения. Этот путь пространственно короток, но преодолевается он с большим трудом, так как клеточные мембраны препятствуют передвижению воды. Этот ток снабжает водой все органы и ткани, приводя их в состояние тургора. Восходящий ток воды захватывает мин. ионы, поглощенные корнем, переносит их и тем самым облегчает распределение (но не поглощение!) по растению.
Концевые двигатели водного тока.
Для того чтобы вода двигалась по растению (и не просто двигалась, а поднималась вверх), необходимы затраты энергии, точки приложения которой располагаются по концам тока, вследствие чего они получили название концевых двигателей.
Нижний концевой двигатель, или корневое давление. Его роль проявляется в основном при активном поглощении — нагнетании воды. При участии актомиозиноподобных сократительных белков он не только подает воду в корневую систему, но и проталкивает ее дальше в сосуды корня и вверх по стеблю. Нагнетание воды — активный энергозависимый процесс, который сильнее всего выражен в коре корня. Сила, развиваемая концевым двигателем, невелика, она может обеспечить подъем воды на высоту не более 1 м., т. е. достаточна для травянистых растений и небольших кустарников. Верхний концевой двигатель, или присасывающая сила транспирации. При постоянном испарении воды в листьях растений разбивается сосущая сила, отсасывающая воду от ближайших клеток и передающаяся др. кл., по к-ым передвигается вода, вплоть до сосудов. В сосудах нет цитоплазмы, поэтому нет и осмотического давления, и всасывание жидкости происходит при участии всей величины сосущей силы. Она позволяет поднять воду на несколько метров. Этой силы дост. для обеспечения водой кустарников и небольших деревьев.
Поднятие воды по стволу
Многие древесные раст. имеют большую протяженность ствола. У таких растений на помощь приходят силы сцепления между молекулами воды, которые очень велики и могут достигать 30 — 35 МПа. Этой силы достаточно для того, чтобы поднять воду на 1 — 2 км, что значительно превышает высоту любого дерева. Силы сцепления молекул воды действуют только при определенных условиях: водные струи в сосудах должны идти непрерывно, без пузырьков воздуха. Если же в них попадает в-х, передвижение воды прерывается. Непрерывность водных струй в сосудах достигается их сильным натяжением и плотным прилеганием к стенкам сосудов. Водные струи оказываются как бы висящими, они удерживаются в подвешенном состоянии сосущей силой трансп. и снизу поддерживаются силой корневого Р. При сильном натяжении водных струй пр-ит натяжение и самих сосудов, при этом у последних наблюдается увелич. длины и уменьшение поперечного сечения (подобно эластичной резине). Это создает предпосылки к смыканию стенок сосудов, что привело бы к их закрыванию и прекращению водного тока. Однако этого не пр-ит, т. к. сосуды имеют внутренние утолщения стенок — кольчатые, спиральные, лестничные, к-ые наподобие каркаса растягивают их и предохр. от смыкания. Величина его незначительна — всего 20 см воды на 1 см2 площади сосудов. Однако и этого достаточно при условии не очень сильного напряжения атмосферных факторов.
Физиологическая значимость процессов минерального питания, коэффициент накопления, органогены, определение макро- и микроэлементов, их представители. Первичные и вторичные макроэлементы. Тройное правило Арнона.
Растения могут поглощать и перемещать неорганические и органические вещества. Раньше считали, что для роста растений (фотосинтезирующих автотрофов) необходимы только минеральные элементы. Однако в последние годы показано, что многие водоросли требуют витамины В12, а также веществ, содержащих кобальт и т. д.
Для наземных растений основным источником воды и минеральных веществ служит почва, не менее важна и атмосфера, которая обеспечивает фотосинтезирующие растения СО2, и, в свою очередь, сама атмосфера поглощает О2 и воду, выделяемую при транспирации.
В системе растение-почва-атмосфера происходит значительный круговорот веществ, особенно воды и СО2.
Концентрация минеральных веществ в самих растениях в большинстве случаев мало связана с их концентрацией в среде. Например, соотношение K+/Na+ в растениях обычно составляет 5–20, тогда как в среде, как правило, больше Na+. С другой стороны, элементы, содержащиеся в окружающей среде в низких концентрациях, могут накапливаться в растениях в значительном количестве.
Эта способность растений характеризуется так называемым коэффициентом накопления:
 |
Еще раз отметим, что ни присутствие, ни концентрация минеральных элементов в самих растениях не является критерием необходимости. Хорошо зарекомендовало себя в этом отношении (относительно необходимости) так называемое тройное правило Арнона, которое было предложено еще в 1939 году. Элемент признается необходимым в случае, когда: 1. Растение без него не может закончить свой жизненный цикл; 2. Другой элемент не может заменить функцию изучаемого элемента; 3. Элемент непосредственно включен в метаболизм растения.
Питательные элементы, которые используются растениями из субстрата в больших количествах (содержание их в золе 10 –1 –10 –2 %), называют макроэлементами, а необходимые в значительно меньшем количестве (содержание 10 –3 % и ниже) – микроэлементами.
Вода и СО2 рассматриваются отдельно, но в этом контексте необходимо считать их макроэлементами. К макроэлементам относятся K, Ca, Mg, N, P, S (Н2О и СО2), для галофитов следует добавить Na и Cl.
Микроэлементы: Mn, Zn, Cu, Cl, B, Mo, сюда же относится и Fe, который располагается на границе макро- и микроэлементов.
Водорослям необходим один или несколько из следующих элементов: Со, Si, I, V. Некоторым высшим растениям необходимы селен, кремний.
Кроме необходимых элементов для роста, в оптимальных пределах должны быть и другие факторы: рН, температура, обеспечение О2, градиент водного потенциала, а для зеленых растений – освещенность соответствующей интенсивности и протяженности.
Пассивный транспорт – перемещение веществ путем диффузии по градиенту электрохимического потенциала без затраты энергии (простая и, в какой-то мере, облегченная диффузия).
Активный транспорт – перемещение веществ против градиента электрохимического потенциала с затратой метаболической энергии, как правило в форме АТФ или редокс-цепей.
Для того чтобы понять механизмы трансмембранного переноса элементов минерального питания, остановимся на рассмотрении некоторых физико-химических закономерностях, определяющих движение ионов в растворе и мембране. Начнем с процессов пассивного транспорта (в частности вспомним законы диффузии).
Согласно первому закону Фика, поток (Ф) прямо пропорционален коэффициенту диффузии D и градиенту концентрации dC/dх в точке х в данный момент времени.
Знак минус в правой части уравнения означает, что, если градиент положителен, т. е. концентрация при увеличении х возрастает, диффузионный поток направлен в противоположную сторону.
Однако при описании диффузии на большие расстояния в непрерывной системе уравнения (5.1) необходимо определить зависимость концентрации не только от расстояния, но и от времени. Поэтому целесообразно преобразовать первый закон Фика в дифференциальные уравнения в частных производных, которые обычно называют вторым законом Фика. В одномерном случае этот закон аналитически выражается следующим образом:
т. е. скорость изменения концентрации пропорциональна второй производной от концентрации на координате х. Из этого закона мы получаем очень важное соотношение:
движение ионов через мембрану обусловлено градиентом электрохимического потенциала.
Обычно мембрана на внутренней стороне отрицательно заряжена по отношению к наружной поверхности. Это приводит к преимущественному поступлению катионов по сравнению с анионами. В этом случае физической движущей силой будет электрохимический потенциал 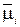 , в величину которого вносят свой вклад химический и электрический потенциалы:
, в величину которого вносят свой вклад химический и электрический потенциалы:
где – электрохимический потенциал i-иона в стандартных условиях; Zi – валентность i-иона; F – число Фарадея; φ – электрический потенциал.
 | (5.10) |
Последнее изменение этой страницы: 2017-07-22; просмотров: 206
Что такое верхний концевой двигатель
Вода и жизнь. Распространение воды в природе. Качественный количественный состав, круговорот, возраст и генезис воды. Изотопный состав, тяжелая вода. Чистая и сверхчистая вода. Способы ее опреснения и очистки. Эффект обезвоживания веществ. Особые свойства жидкой воды — реальные и вымышленные.
Физические свойства воды. Особенности строения молекул, водородные связи и модели структуры жидкой воды. Коэффициент самодиффузии как показатель микроподвижности молекул.
Водные растворы. Взаимодействие водной структуры с растворенным веществом.
Различные виды гидратации: (гидрофильная и гидрофобная, ионная и электронейтральная).
Растворы макромолекул. Вода свободная и связанная, капиллярная, коллоидная, иммобилизованная. Вода в биологических структурах. Клатратные комплексы и их свойства. Соотношение различных фракций воды в растении и их физиологическое значение. Методы оценки состояния воды в биологических объектах: ЯМР-, ИК- и диэлектрическая спектроскопия.
Макроперемещения молекул воды в растении. Факторы, влияющие на этот процесс.
Водный потенциал как обобщенный энергетический показатель состояния воды в биологических системах. Термодинамические показатели состояния воды и ее растворов. Понятие об энергии. Энергия внутренняя, тепловая, свободная. Законы термодинамики. Химический потенциал. Зависимость химического потенциала от концентрации. Активность воды и способы ее определения. Ионная сила раствора. Полный потенциал системы и его составляющие. Значение, способы и единицы выражения химического, полного и водного потенциалов. Диффузия. Движущая сила этого процесса. Законы А.Фика. Осмотический потенциал водных растворов, выводы Вант-Гоффа. Связь водного потенциала с полным, осмотическим потенциалами и с сосущей силой клетки.
Давление в жидкости. Гидростатическое, осмотическое и тургорное давление. Тургор и плазмолиз в растительных клетках. Методы определения осмотического и водного потенциалов биологических объектов. Осмотики, используемые при исследованиях и предъявляемые к ним требования. Коэффициент отражения биологических мембран.
Потоки воды в целом растении. Большой круг циркуляции воды в системе почва — растение-атмосфера. Движущие силы на этом пути. Радиальный транспорт воды в корне. Анатомические особенности строения корня. Нижний концевой двигатель растений. Осмотическая и метаболическая составляющие корневого давления. Представления о пассивном и активном участии растения в процессах водообмена. Верхний концевой двигатель растений. Транспирация. Поток воды на пути ксилемные окончания-воздух. Способы регуляции этого потока. Строение и функциональне особенности устиц. Движение воды по внутриннему апопласту растений. Поверхностное натяжение и капилярные силы. Значение когезии и адгезии для транспорта воды. Анатомические особенности строения проводящих путей ксилемы.
Малый круг циркуляции воды в растении. Поток через флоэму и силы его регулирующие. Строение проводящей системы флоэмы. Некоторые модели флоэмного двигателя и его роль в жизни растений. Влияние внешних и внутренних факторов на водообмен растений. Эволюция проводящих систем у растений. Особенности растений различных экологических групп и пути их адаптации к условиям водосна.
Виды транспирации, верхний концевой двигатель, устройство и работа устьичного механизма.
Транспирацией называется физиологическое испарение воды растениями. Основным органом транспирации является лист.
Основные структуры участвующие в процессе транспирации это:
· Устьица – поры, через которые диффундирует вода, испаряющаяся в поверхности клеток. Устьица находятся в эпидермисе листьев и стеблей (для травянистых растений). Через устьица испаряется до 90% воды. Устьичная транспирация.
· Кутикула – воскоподобный слой, покрывающий эпидермис листьев и стеблей. Через него проходит вода, испаряющаяся с наружных стенок клеток эпидермиса. Через кутикулу испаряется около 10% воды, в зависимости от толщины кутикулы. Кутикулярная транспирация.
· Чечевички: обычно их роль невелика, но у листопадных деревьев после опадения листьев через них испаряется основная масса воды.
Испарение воды в результате транспирации очень велико. Травянистые растения такие как подсолнечник или кукуруза могут испарять за сутки 1-2 литра воды, а зрелые деревья, например старый дуб более 600 литров в сутки.
В результате испарения воды клетками листа в них снижается обводненность и повышается сосущая сила. Это приводит к повышению поглощения воды клетками листа из сосудов ксилемы и передвижению воды по ксилеме из корней к листу (т.е. наблюдается присасывающее действие листьев).
Т.о. верхний концевой двигатель создается и поддерживается высокой сосущей силой транспирирующих клеток листовой паренхимы.
Рассмотрим движение воды по листу и механизм транспирации.

Вода поступает в лист по сосудам ксилемы. В листе проводящий пучок ветвится на сеть уменьшающихся по диаметру жилок. Сеть сосудов настолько густа, сто ни одна клетка листа не отделена от сосуда более чем двумя рядами клеток. Каждая жилка оканчивается трахеидой или мелким сосудом с мало лигнифицированными стенками. Поэтому вода легко поступает в клетки листа.
Далее вода может идти тремя путями: по апопласту она движется наиболее быстро, транспорт через симпласт наиболее медленный.
Движение воды через вакуоли представляет собой смешение двух предыдущих путей. В этом случае она проходит как через цитоплазму клеток, так и через апопласт. То есть присутствуют более и менее медленные участки пути.
Вода в листе движется по градиенту водного потенциала, сходному с таковым в корнях с тем лишь отличием, что наибольшее количество воды находится в клетках примыкающих к сосуду, а стало быть, их сосущая сила ниже, чем у расположенных далее.
Все три пути завершаются процессом транспирации. Сам процесс можно разделить на три стадии:
1. Испарение воды с клеточных стенок в межклетники.
2. Диффузия паров воды через устьица наружу.
3. Диффузия паров воды от поверхности листа.
Все листья окружены неподвижным слоем воздуха, толщина которого определяется размером листа и особенностями его строения, например опушением. Пары воды сначала диффундируют через этот слой, а потом уносятся воздушными потоками.
 На рисунке линиями показаны области одинаковой концентрации паров воды. Чем выше градиент водного потенциала, тем ближе расположены эти линии и тем быстрее происходит диффузия. Поэтому имеет место краевой эффект, т.е. диффузия идет быстрее с края устьица, чем по центру. Этим и объясняется, что испарение воды идет быстрее при большом числе малых отверстий, чем при большом отверстии той же общей площади.
На рисунке линиями показаны области одинаковой концентрации паров воды. Чем выше градиент водного потенциала, тем ближе расположены эти линии и тем быстрее происходит диффузия. Поэтому имеет место краевой эффект, т.е. диффузия идет быстрее с края устьица, чем по центру. Этим и объясняется, что испарение воды идет быстрее при большом числе малых отверстий, чем при большом отверстии той же общей площади.
Механизм передвижеия воды из корня в надземные органы. Верхний и нижний концевые двигатели.
Т. к. вода поглощается в осн. корневой системой, возникает необходимость ее передвижения по растению. Этот процесс составляет т. н. восходящий ток. Следует отметить, что это название отражает характер передвижения и его локализацию в растении. Он проходит по мертвым тканям стебля или черешка — сосудам у покрытосеменных и трахеидам у голосеменных. Однако эта локализация не абсолютна: вода способна перемещаться и по флоэмной системе. Передвижение по сосудам идет с большей скоростью (20 см/ч), чем по трахеидам (только 5 см/ч).
Если принять во внимание всю протяженность пути восходящего тока, то его можно будет разбить на 2 не равных по длине участка: Мертвые гистологические элементы в середине проводящего пути — сосуды или трахеиды. Протяженность значительна, но вода проходит по нему сравнительно легко, так как двигается пассивно по мертвым элементам, не испытывая с их стороны значительного сопротивления. Живые клетки корня и листа, находящиеся в начале и в конце пути передвижения. Этот путь пространственно короток, но преодолевается он с большим трудом, так как клеточные мембраны препятствуют передвижению воды. Этот ток снабжает водой все органы и ткани, приводя их в состояние тургора. Восходящий ток воды захватывает мин. ионы, поглощенные корнем, переносит их и тем самым облегчает распределение (но не поглощение!) по растению.
Концевые двигатели водного тока.
Для того чтобы вода двигалась по растению (и не просто двигалась, а поднималась вверх), необходимы затраты энергии, точки приложения которой располагаются по концам тока, вследствие чего они получили название концевых двигателей.
Нижний концевой двигатель, или корневое давление. Его роль проявляется в основном при активном поглощении — нагнетании воды. При участии актомиозиноподобных сократительных белков он не только подает воду в корневую систему, но и проталкивает ее дальше в сосуды корня и вверх по стеблю. Нагнетание воды — активный энергозависимый процесс, который сильнее всего выражен в коре корня. Сила, развиваемая концевым двигателем, невелика, она может обеспечить подъем воды на высоту не более 1 м., т. е. достаточна для травянистых растений и небольших кустарников. Верхний концевой двигатель, или присасывающая сила транспирации. При постоянном испарении воды в листьях растений разбивается сосущая сила, отсасывающая воду от ближайших клеток и передающаяся др. кл., по к-ым передвигается вода, вплоть до сосудов. В сосудах нет цитоплазмы, поэтому нет и осмотического давления, и всасывание жидкости происходит при участии всей величины сосущей силы. Она позволяет поднять воду на несколько метров. Этой силы дост. для обеспечения водой кустарников и небольших деревьев.
Поднятие воды по стволу
Многие древесные раст. имеют большую протяженность ствола. У таких растений на помощь приходят силы сцепления между молекулами воды, которые очень велики и могут достигать 30 — 35 МПа. Этой силы достаточно для того, чтобы поднять воду на 1 — 2 км, что значительно превышает высоту любого дерева. Силы сцепления молекул воды действуют только при определенных условиях: водные струи в сосудах должны идти непрерывно, без пузырьков воздуха. Если же в них попадает в-х, передвижение воды прерывается. Непрерывность водных струй в сосудах достигается их сильным натяжением и плотным прилеганием к стенкам сосудов. Водные струи оказываются как бы висящими, они удерживаются в подвешенном состоянии сосущей силой трансп. и снизу поддерживаются силой корневого Р. При сильном натяжении водных струй пр-ит натяжение и самих сосудов, при этом у последних наблюдается увелич. длины и уменьшение поперечного сечения (подобно эластичной резине). Это создает предпосылки к смыканию стенок сосудов, что привело бы к их закрыванию и прекращению водного тока. Однако этого не пр-ит, т. к. сосуды имеют внутренние утолщения стенок — кольчатые, спиральные, лестничные, к-ые наподобие каркаса растягивают их и предохр. от смыкания. Величина его незначительна — всего 20 см воды на 1 см2 площади сосудов. Однако и этого достаточно при условии не очень сильного напряжения атмосферных факторов.
Физиологическая значимость процессов минерального питания, коэффициент накопления, органогены, определение макро- и микроэлементов, их представители. Первичные и вторичные макроэлементы. Тройное правило Арнона.
Растения могут поглощать и перемещать неорганические и органические вещества. Раньше считали, что для роста растений (фотосинтезирующих автотрофов) необходимы только минеральные элементы. Однако в последние годы показано, что многие водоросли требуют витамины В12, а также веществ, содержащих кобальт и т. д.
Для наземных растений основным источником воды и минеральных веществ служит почва, не менее важна и атмосфера, которая обеспечивает фотосинтезирующие растения СО2, и, в свою очередь, сама атмосфера поглощает О2 и воду, выделяемую при транспирации.
В системе растение-почва-атмосфера происходит значительный круговорот веществ, особенно воды и СО2.
Концентрация минеральных веществ в самих растениях в большинстве случаев мало связана с их концентрацией в среде. Например, соотношение K+/Na+ в растениях обычно составляет 5–20, тогда как в среде, как правило, больше Na+. С другой стороны, элементы, содержащиеся в окружающей среде в низких концентрациях, могут накапливаться в растениях в значительном количестве.
Эта способность растений характеризуется так называемым коэффициентом накопления:
 |
Еще раз отметим, что ни присутствие, ни концентрация минеральных элементов в самих растениях не является критерием необходимости. Хорошо зарекомендовало себя в этом отношении (относительно необходимости) так называемое тройное правило Арнона, которое было предложено еще в 1939 году. Элемент признается необходимым в случае, когда: 1. Растение без него не может закончить свой жизненный цикл; 2. Другой элемент не может заменить функцию изучаемого элемента; 3. Элемент непосредственно включен в метаболизм растения.
Питательные элементы, которые используются растениями из субстрата в больших количествах (содержание их в золе 10 –1 –10 –2 %), называют макроэлементами, а необходимые в значительно меньшем количестве (содержание 10 –3 % и ниже) – микроэлементами.
Вода и СО2 рассматриваются отдельно, но в этом контексте необходимо считать их макроэлементами. К макроэлементам относятся K, Ca, Mg, N, P, S (Н2О и СО2), для галофитов следует добавить Na и Cl.
Микроэлементы: Mn, Zn, Cu, Cl, B, Mo, сюда же относится и Fe, который располагается на границе макро- и микроэлементов.
Водорослям необходим один или несколько из следующих элементов: Со, Si, I, V. Некоторым высшим растениям необходимы селен, кремний.
Кроме необходимых элементов для роста, в оптимальных пределах должны быть и другие факторы: рН, температура, обеспечение О2, градиент водного потенциала, а для зеленых растений – освещенность соответствующей интенсивности и протяженности.
Пассивный транспорт – перемещение веществ путем диффузии по градиенту электрохимического потенциала без затраты энергии (простая и, в какой-то мере, облегченная диффузия).
Активный транспорт – перемещение веществ против градиента электрохимического потенциала с затратой метаболической энергии, как правило в форме АТФ или редокс-цепей.
Для того чтобы понять механизмы трансмембранного переноса элементов минерального питания, остановимся на рассмотрении некоторых физико-химических закономерностях, определяющих движение ионов в растворе и мембране. Начнем с процессов пассивного транспорта (в частности вспомним законы диффузии).
Согласно первому закону Фика, поток (Ф) прямо пропорционален коэффициенту диффузии D и градиенту концентрации dC/dх в точке х в данный момент времени.
Знак минус в правой части уравнения означает, что, если градиент положителен, т. е. концентрация при увеличении х возрастает, диффузионный поток направлен в противоположную сторону.
Однако при описании диффузии на большие расстояния в непрерывной системе уравнения (5.1) необходимо определить зависимость концентрации не только от расстояния, но и от времени. Поэтому целесообразно преобразовать первый закон Фика в дифференциальные уравнения в частных производных, которые обычно называют вторым законом Фика. В одномерном случае этот закон аналитически выражается следующим образом:
т. е. скорость изменения концентрации пропорциональна второй производной от концентрации на координате х. Из этого закона мы получаем очень важное соотношение:
движение ионов через мембрану обусловлено градиентом электрохимического потенциала.
Обычно мембрана на внутренней стороне отрицательно заряжена по отношению к наружной поверхности. Это приводит к преимущественному поступлению катионов по сравнению с анионами. В этом случае физической движущей силой будет электрохимический потенциал  , в величину которого вносят свой вклад химический и электрический потенциалы:
, в величину которого вносят свой вклад химический и электрический потенциалы:
где – электрохимический потенциал i-иона в стандартных условиях; Zi – валентность i-иона; F – число Фарадея; φ – электрический потенциал.